Article Type: Research Article, Volume 2 Issue 1
*Corresponding author: Bo Pan
Hunan Provincial Hospital of Integrated Traditional Chinese and Western Medicine, Hunan 410006, China.
Email: 15675190855@163.com
Received: Jan 18, 2025 Accepted: Feb 21, 2025 Published: Feb 28, 2025
Citation: Wang W, Li L, Yang B, Shen S, Pan B, et al. Serum metabolites in the modulation of inflammatory factors on hepatic cancer: Mendelian randomization and mediation analysis. Ann Case Rep Med Images. 2025; 2(1): 1016.
Copyright: Pan B et al. © All rights are reserved
Objective:To explore the causal relationships between inflammatory factors and hepatic cancer and identify the role of serum metabolites as potential mediators.
Methods:In this study, we employed two-sample Mendelian Randomization (MR) analysis based on publicly available large-scale Genome-Wide Association Study (GWAS) data to investigate the causal relationships between 91 inflammatory factors, 1400 serum metabolites, and hepatic Cancer. The Inverse Variance-Weighted (IVW) MR was used as the primary analytical method, supplemented by additional sensitivity tests to assess the validity of causal relationships. Furthermore, we employed a two-step mediation MR to identify potential mediating serum metabolites and evaluate their mediating effects.
Results:After adjusting for heterogeneity and horizontal pleiotropy, MR analysis identified 6 inflammatory factors such as C-C motif chemokine 19 levels, Fms-related tyrosine kinase 3 ligand levels, and Interleukin-13 levels, which were causally related to hepatic Cancer. In the causal relationship study between serum metabolites and hepatic Cancer, we identified 73 serum metabolites such as S-allylcysteine levels, (1-enyl-palmitoyl)-2-linoleoyl-GPE (p-16:0/18:2) levels, and Caffeic acid sulfate levels that were causally related to hepatic Cancer. Mediation analysis in MR suggested that the inflammatory factor Interleukin-13 levels and Interleukin-4 levels could respectively increase the levels of serum metabolite Linolenate [alpha or gamma; (18:3n3 or 6)] levels and Adenosine 5’-Diphosphate (ADP) to choline ratio, thereby increasing the risk of hepatic Cancer, with a mediation effect of 24.6% and 6.5%, respectively. Conversely, the inflammatory factor Monocyte chemoattractant protein-3 levels could decrease the levels of serum metabolite Sphingomyelin (d18:0/20:0, d16:0/22:0) levels, thereby reducing the risk of hepatic Cancer, with a mediation effect of 5.4%.
Conclusion:This study demonstrates significant causal relationships between certain inflammatory factors, serum metabolites, and hepatic Cancer. Three serum metabolites were found to have mediating effects in the influence of inflammatory factors on the risk of hepatic Cancer. This finding provides a new perspective for understanding the role of inflammatory factors in the development of hepatic Cancer and offers potential targets for the prevention and treatment of related diseases.
Keywords: Hepatic cancer; Inflammatory factors; Serum metabolites; Mendelian randomization; Mediation analysis
Hepatic Cancer stands among the most commonly occurring neoplasms worldwide and ranks as a leading cause of cancerrelated mortality. Inflammatory factors are intricately linked with the progression of hepatic cancer and play a pivotal role in its developmental stages. Serum metabolites, the byproducts of the body’s metabolic processes, display an association with the incidence and evolution of hepatic cancer.
Inflammatory factors, as essential regulatory molecules, exert complex effects during the progression of hepatic cancer. The inflammatory milieu can activate tumor-associated signaling pathways, promoting tumor proliferation, invasion, and metastasis. Additionally, inflammatory factors can facilitate the development of hepatic cancer by activating immune cells, stimulating angiogenesis, and altering the cell cycle. Serum metabolites represent a widespread class of small organic molecules within the body, with levels influenced by genetic factors, environmental conditions, and physiological states. Studies have indicated that the levels of serum metabolites such as amino acids, lipid metabolism disorders, and cholesterol are closely associated with the occurrence and progression of hepatic cancer [1]. Moreover, there is a potential association between serum metabolites and inflammatory factors in hepatic cancer. Research has shown that inflammatory factors can induce oxidative stress, and serum Reactive Oxygen Species (ROS) metabolites serve as reliable indicators of circulating ROS, which are correlated with the induced progression of hepatic cancer [2].
However, the causal relationships between inflammatory factors, serum metabolites, and hepatic cancer remain unclear. This study utilizes publicly available large-scale GWAS data to conduct a two-sample Mendelian Randomization (MR) mediation analysis, aiming to investigate the causal relationships and mediating effects among inflammatory factors, serum metabolites, and hepatic cancer. By further elucidating the mechanisms underlying the development of hepatic cancer, this research provides new theoretical insights for the prevention and treatment of hepatic cancer.
Study design
We conducted a two-sample MR analysis to investigate the causal relationships among 91 inflammatory factors, 1400 serum metabolites, and hepatic cancer. Additionally, we employed serum metabolites as intermediate factors and conducted MR mediation analysis to investigate whether serum metabolites mediate the relationship between inflammatory factors and the risk of hepatic cancer incidence. MR harnesses genetic variants as proxies for risk factors, necessitating that Instrument Variables (IVs) fulfill three cardinal presumptions for valid causal inference: (1) genetic variants are robustly associated with exposure; (2) genetic variants are uncorrelated with any potential confounders of the exposure-outcome relationship (i.e., no horizontal pleiotropy); (3) genetic variants influence the outcome solely through the exposure, not by any alternative pathways. Following the two-sample MR analysis, this study conducted MR mediation analysis on inflammatory factors to decompose the direct and indirect effects of inflammatory factors and serum metabolites on hepatic cancer. The overall design of this study is illustrated in Figure 1.
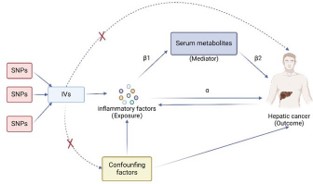
Figure 1: Study design of two-sample MR and MR mediation analysis.
Source of exposure factors GWAS Data
We selected Single Nucleotide Polymorphisms (SNPs) significantly associated with inflammatory factors and serum metabolites as IVs. The complete summary statistics data for inflammatory factors and serum metabolites GWASs can be obtained from the catalog database (https://www.ebi.ac.uk/gwas/pub- lications/37563310) GWAS catalog [3-5]. A total of 91 inflammatory factors (GCST90274758 to GCST90274848) and 1,400 serum metabolites (GCST90199621 to GCST90201020) were included. The inflammatory factors are derived from the latest comprehensive GWAS summary data, which include 14,824 Europeans from 11 cohorts, involving 91 systemic inflammation regulatory factors measured using the Olink Target inflammation panel and subjected to whole-genome pQTL localization. The serum metabolites encompass 1,091 serum metabolites and 309 metabolite ratios, derived from 8,299 individuals in the Canadian Longitudinal Study on Aging (CLSA) cohort.
Source of hepatic cancer GWAS data
We downloaded the GWAS data for hepatic cancer (GWAS ID: ebi-a-GCST90018858) reported in the IEU Open GWAS (https://gwas.mrcieu.ac.uk/) database, comprising 475,638 samples and 24,194,938 SNPs. The study population consisted of Europeans, and ethical approval was not required as the data were obtained from public databases.
Selection of IVs
We selected genetic variants (SNPs) exhibiting strong association with the exposure factors as IVs to examine the causal relationship between exposure factors and outcomes. GWAS data sets were utilized to extract SNPs corresponding to inflammatory factors, with SNPs as exposure factors required to meet a significance threshold lower than the genome-wide statistical significance threshold (5.0×10−8). If the number of IVs obtained was <5, we lowered the threshold to P<5×10−6 based on methods from previous MR studies [6]. Additionally, to mitigate bias caused by Linkage Disequilibrium (LD), we restricted SNP clumping to clump_kb >10,000 and clump_r2 <0.001, selecting only the SNP with the strongest effect on outcomes as the genetic instrument. The calculation of the R2 for each SNP in the IVs involved the formula: R2=2×(1-MAF)×MAF×β2, where R2 represents the degree to which the instrumental variable explains the exposure, MAF represents the minor allele frequency, and β represents the allele effect size. Subsequently, we calculated the F value as F=R2×(n-k-1)/[k×(1R2)], where n represents the total sample size and k represents the number of instrumental variables. Finally, weak instrumental variables bias was removed, and only SNPs with F>10 were considered for inclusion in the MR analysis.
Statistical analysis
To assess the causal relationships between inflammatory factors, serum metabolites, and hepatic cancer, we used selected SNPs as IVs and conducted analyses using the “Mendelian Randomization” package [7]. We primarily employed three methods, namely, the conventional fixed-effects Inverse Variance-Weighted (IVW) [8], the Weighted Median (WM) [9], and the MR-Egger method to estimate the causal effects of exposure on outcomes, with forest plots illustrating the analytical results. Cochran’s Q test was used for heterogeneity testing, with P<0.05 indicating heterogeneity. Where heterogeneity was observed with P<0.05, the random-effects IVW method substituted the fixed-effects model to correct for the heterogeneity [8,10]. The IVW method is reported to be marginally superior under certain conditions but presumes all IVs are valid [9].
Therefore, while the IVW method serves as the primary MR analysis method, we also used the weighted median method [9] and leave-one-out sensitivity testing [11] to evaluate the robustness of causal effect estimates after removing one SNP at a time, as the weighted median method is more robust to outliers. MR-Egger regression was applied for intercept testing to assess horizontal pleiotropy [12,13]. A p-value<0.05 indicates the presence of horizontal pleiotropy, and the remaining SNP list after removing horizontally pleiotropic SNPs was used for subsequent MR analysis. Additionally, scatter plots and funnel plots were used for visualization. The scatter plot demonstrates that the results are not influenced by outliers, while the funnel plot demonstrates the robustness of the correlation and absence of heterogeneity. All statistical analyses were performed using R 4.2.3 (http:// www.Rproject.org), R studio software, and R packages, with a significance level of α=0.05.
Following the completion of the two-sample MR analysis, we employed the Two-Step Mendelian Randomization (TSMR) method to decompose the direct and indirect effects of inflammatory factors and serum metabolites on hepatic cancer [14]. TSMR assumes no interaction between exposure and mediator variables. In addition to the total effect estimate of inflammatory factors on hepatic cancer obtained from the univariable MR analysis (α), two additional estimates were computed: (1) the causal effect of inflammatory factors on serum metabolites (β1); (2) the causal effect of serum metabolites on hepatic cancer (β2). The mediation effect, which refers to the causal effect of inflammatory factors on hepatic cancer via mediators, can then be estimated using the product of coefficients method (β1*β2). Thus, the proportion mediated could be calculated as “mediation effect/total effect” ([β1*β2]/ α).
Causal relationship between inflammatory factors and hepatic cancer
To investigate the causal impact of inflammatory factors on hepatic cancer, MR analysis was conducted using the IVW method, Weighted median method, and MR-Egger as the main analytical approaches. GWAS datasets were utilized to extract SNPs of inflammatory factors as IVs, with these SNPs as exposure factors needing to reach a significance threshold of less than the genome-wide statistical threshold (5×10−8). However, after selecting SNPs, only a small number of inflammatory factors were chosen as IVs, prompting us to lower the threshold to P<5×10−6 to discover more comprehensive causal associations between inflammatory factors and hepatic cancer.
Following heterogeneity testing, sensitivity analysis, and horizontal pleiotropy testing (see specific results in section 3.4), IVW method identified a total of 6 inflammatory factors with a causal relationship with hepatic cancer (Figure 2A). Among them, 4 inflammatory factors were positively associated with an increased risk of hepatic cancer, while 2 inflammatory factors were negatively associated with the risk of hepatic cancer (Figure 2). We found that C-C motif chemokine 19 levels (GCST90274765) (IVW: OR=1.258, 95% CI=1.090–1.452, P=1.677×10-3; Weighted median: OR=1.260, 95% CI=1.068– 1.486, P=6.15×10-3; MR Egger: OR=1.327, 95% CI=1.073–1.642, P=0.018) (Figure 3) and Fms-related tyrosine kinase 3 ligand levels (GCST90274791) (IVW: OR=1.290, 95% CI=1.045–1.593, P=0.018; Weighted median: OR=1.405, 95% CI=1.052–1.876, P=0.021; MR Egger: OR=1.806, 95% CI=1.093–2.981, P=0.029) were causally associated with hepatic cancer, acting as risk factors, with all three analytical methods demonstrating statistical significance. Similarly, Interleukin-13 levels (GCST90274799) and Interleukin-4 levels (GCST90274813) were also risk factors for hepatic cancer.
On the other hand, Interleukin-10 levels (GCST90274795) (IVW: OR=0.638, 95% CI=0.453–0.899, P=0.010) showed a causal relationship with hepatic cancer according to the IVW method, serving as a protective factor. Monocyte chemoattractant protein-3 levels (GCST90274823) (IVW: OR=0.742, 95% CI=0.556–0.989, P=0.042; Weighted median: OR=0.666, 95% CI=0.465–0.954, P=0.027), as indicated by IVW and Weighted median methods, showed a causal relationship with hepatic cancer, also acting as a protective factor. The MR results from the IVW, Weighted median, and MR-Egger methods were visualized using forest plots (Figure 2B).
Furthermore, we conducted reverse MR analysis on the 6 selected inflammatory factors with hepatic cancer, with IVW as the main analytical method. The results showed no causal relationship between hepatic cancer and the 6 inflammatory factors, with P>0.05 (Table 1).

Figure 2: Forest plot of the causal relationship between inflammatory factors and hepatic cancer (A) Forest plot of IVW method MR analysis; (B) Forest plot of IVW, Weighted median, and MR-Egger method MR analysis).
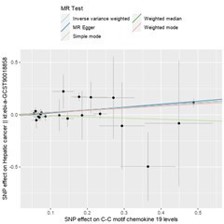
Figure 3: Comparison of the strongest inflammatory factors associated with hepatic cancer using different MR methods.
| Exposure | ID outcome | Outcome | Beta | SE | P value | OR (95%CI) |
|---|---|---|---|---|---|---|
| Hepatic cancer | GCST90274765 | C-C motif chemokine 19 levels | 0.053 | 0.043 | 0.219 | 1.054(0.969-1.147) |
| Hepatic cancer | GCST90274791 | Fms-related tyrosine kinase 3 ligand levels | 0.036 | 0.026 | 0.164 | 1.037(0.985-1.091) |
| Hepatic cancer | GCST90274795 | Interleukin-10 levels | -0.003 | 0.056 | 0.964 | 0.997(0.893-1.114) |
| Hepatic cancer | GCST90274799 | Interleukin-13 levels | -0.027 | 0.024 | 0.262 | 0.974(0.930-1.020) |
| Hepatic cancer | GCST90274813 | Interleukin-4 levels | -0.011 | 0.024 | 0.636 | 0.989(0.944-1.036) |
| Hepatic cancer | GCST90274823 | Monocyte chemoattractant protein-3 levels | 0.013 | 0.012 | 0.289 | 1.013(1.016-1.114) |
Causal relationship between serum metabolites and hepatic cancer
Following heterogeneity testing, sensitivity analysis, and horizontal pleiotropy testing, GWAS datasets were utilized to extract SNPs of serum metabolites as IVs, with selected SNPs as exposure factors needing to reach a significance threshold of less than the genome-wide statistical threshold (5×10−6). Using the IVW method, we identified a total of 73 serum metabolites with a causal relationship with the occurrence of hepatic cancer (Figure 4, displaying the top 30 ranked serum metabolites). Among them, 32 serum metabolites were negatively associated with the risk of hepatic cancer, while the remaining 41 serum metabolites were positively associated with the risk of hepatic cancer.
Among the identified serum metabolites, S-allylcysteine levels (GCST90199948) (IVW: OR=0.685, 95% CI=0.537-0.874, P=0.002; Weighted median: OR=0.707, 95% CI=0.510-0.981, P=0.038), (1-enyl-palmitoyl)-2-linoleoyl-GPE (p-16:0/18:2) levels (GCST902000751) (IVW: OR=0.756, 95% CI=0.613-0.932, P=0.008; Weighted median: OR=0.767, 95% CI=0.592-0.993, P=0.044), and Caffeic acid sulfate levels (GCST90200093) (IVW: OR=0.696, 95% CI=0.522-0.929, P=0.014; Weighted median: OR=0.783, 95% CI=0.633-0.969, P=0.025) exhibited significant causal relationships with hepatic cancer in both MR analysis methods, with p<0.05, acting as protective factors for hepatic cancer.
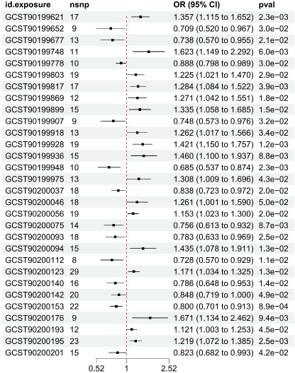
Figure 4: Forest plot of the causal relationship between serum metabolites and hepatic cancer (Forest plot of IVW method MR analysis).
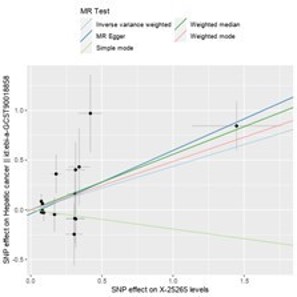
Figure 5: Comparison of different MR methods for the strongest serum metabolites associated with hepatic cancer.
Conversely, X-25265 levels (GCST90200658) (IVW: OR=1.549, 95% CI=1.170-2.051, P=0.002; Weighted median: OR=1.747, 95% CI=1.191-2.563, P=0.004; MR Egger: OR=1.888, 95% CI=1.304-2.733, P=0.005) (Figure 5), Phosphate to mannose ratio (GCST90200769) (IVW: OR=1.265, 95% CI=1.053-1.520, P=0.012; Weighted median: OR=1.407, 95% CI=1.092-1.813, P=0.008; MR Egger: OR=1.769, 95% CI=1.112-2.813, P=0.037), and Glucose-to-mannose ratio (GCST90200953) (IVW: OR=1.202, 95% CI=1.008-1.432, P=0.040; Weighted median: OR=1.320, 95% CI=1.084-1.608, P=0.006; MR Egger: OR=1.684, 95% CI=1.159-2.447, P=0.023) exhibited significant causal relationships with hepatic cancer in all three MR analysis methods, with p<0.05, acting as risk factors for hepatic cancer.
Assessment of result reliability
Heterogeneity testing: Cochran’s Q test conducted for MREgger and IVW methods indicated that in the MR analysis of serum metabolites and hepatic cancer, serum metabolite 1-stearoyl-2-docosahexaenoyl-gpc (18:0/22:6) levels (GCST90200046) had p<0.05, indicating heterogeneity. Consequently, the random-effects IVW method supersedes the fixed-effects IVW to adjust for this heterogeneity. In contrast, in the MR analysis of inflammatory markers associated with hepatic cancer, where p>0.05, no evidence of heterogeneity is observed.
Robustness assessment: Robustness analysis was conducted during the selection of SNPs as instrumental variables using the weighted median method and leave-one-out sensitivity tests, demonstrating the reliability of the MR results. Specifically, in the context of MR analysis between serum metabolites and hepatic cancer, the leave-one-out sensitivity test revealed that SNP rs1260326 might significantly influence the results of Glucose-to-mannose ratio (GCST90200953), while SNP rs57298804 might significantly affect the results of Arachidonate (20:4n6) to paraxanthine ratio (GCST90200977).
Horizontal pleiotropy testing: Intercept testing using MR-Egger regression was conducted to evaluate horizontal pleiotropy. The results indicated that in the MR analysis of inflammatory factors and serum metabolites with hepatic cancer, all p-values were >0.05, suggesting minimal potential influence of genetic pleiotropy on the MR analysis results.
Mendelian randomization mediation analysis
We conducted MR mediation analysis using serum metabolites as the mediators between inflammatory factors and hepatic cancer. Among the 73 serum metabolites causally associated with hepatic cancer, we identified 5 inflammatory factors with causal relationships significantly associated with these 73 serum metabolites. Specifically, three serum metabolites exhibited mediating effects in the impact of inflammatory factors on the risk of hepatic cancer (Figure 6).
Prior to removing the mediator variables, correlations were observed between Interleukin-13 levels, Monocyte chemoattractant protein-3 levels, Interleukin-4 levels and hepatic cancer. However, after removing the mediators, the p-values for the correlations between Interleukin-13 levels and hepatic cancer were 0.091 (P>0.05), for Monocyte chemoattractant protein-3 levels and hepatic cancer were 0.07 (P>0.05), and for Interleukin-4 levels and hepatic cancer were 0.089 (P>0.05), indicating no statistical significance. This suggests that the inflammatory factors Interleukin-13 levels, Monocyte chemoattractant protein-3 levels and Interleukin-4 levels do not have a direct causal effect on hepatic cancer, while the serum metabolites Linolenate [alpha or gamma; (18:3n3 or 6)] levels, Sphingomyelin (d18:0/20:0, d16:0/22:0) levels and Adenosine 5\’-Diphosphate (ADP) to choline ratio may serve as three mediating factors. We found that the inflammatory factor Interleukin-13 levels and Interleukin-4 levels can respectively increase the levels of serum metabolites Linolenate [alpha or gamma; (18:3n3 or 6)] levels and Adenosine 5’-Diphosphate (ADP) to choline ratio, thereby increasing the risk of hepatic cancer occurrence, while the inflammatory factor Monocyte chemoattractant protein-3 levels can decrease the levels of serum metabolites Sphingomyelin (d18:0/20:0, d16:0/22:0) levels, consequently reducing the risk of hepatic cancer occurrence. Specifically, the inflammatory factor Interleukin-13 levels mediate 24.6% of the effect of inflammatory factors on the risk of hepatic cancer through serum metabolites Linolenate [alpha or gamma; (18:3n3 or 6)] levels (α=0.309, β1=0.158, β2=0.484, mediating effect=0.076, direct effect=0.232, proportion mediated=0.246). The inflammatory factor Monocyte chemoattractant protein-3 levels mediate 5.4% of the effect of inflammatory factors on the risk of hepatic cancer through serum metabolites Sphingomyelin (d18:0/20:0, d16:0/22:0) levels (α=-0.299, β1=-0.102, β2=0.158, mediating effect=-0.016, direct effect=-0.283, proportion mediated=0.054). The inflammatory factor Interleukin-4 levels mediate 6.5% of the effect of inflammatory factors on the risk of hepatic cancer through serum metabolites Adenosine 5\’-Diphosphate (ADP) to choline ratio (α=0.309, β1=0.216, β2=0.094, mediating effect=0.020, direct effect=0.288, proportion mediated=0.065) (Tables 2, 3 & 4).

Figure 6: Forest Plot Before and After Removing Mediator Variables.
| Removing mediator effect | Exposure | Outcome | Nsnp | Beta | SE | P value | OR (95%CI) |
|---|---|---|---|---|---|---|---|
| Interleukin-13 levels | Linolenate [alpha or gamma; (18:3n3 or 6)] levels | 11 | 0.158 | 0.072 | 0.029 | 1.171(1.0161.349) | |
| Linolenate [alpha or gamma; (18:3n3 or 6)] levels | Hepatic cancer | 11 | 0.484 | 0.176 | 0.006 | 1.623(1.1492.292) | |
| Before | Interleukin-13 levels | Hepatic cancer | 13 | 0.309 | 0.127 | 0.015 | 1.362(1.0631.745) |
| After | Interleukin-13 levels | Hepatic cancer | 13 | 0.232 | 0.127 | 0.091 | 1.262(1.0621.499) |
| Removing mediator effect | Exposure | Outcome | Nsnp | Beta | SE | P value | OR (95%CI) |
|---|---|---|---|---|---|---|---|
| Monocyte chemoattractant protein-3 levels | Sphingomyelin (d18:0/20:0, d16:0/22:0) levels | 17 | -0.102 | 0.047 | 0.032 | 0.903(0.8230.991) | |
| Sphingomyelin (d18:0/20:0, d16:0/22:0) levels | Hepatic cancer | 29 | 0.158 | 0.063 | 0.013 | 1.171(1.0341.325) | |
| Before | Monocyte chemoattractant protein-3 levels | Hepatic cancer | 19 | -0.299 | 0.147 | 0.042 | 0.742(0.5560.989) |
| After | Monocyte chemoattractant protein-3 levels | Hepatic cancer | 19 | -0.283 | 0.147 | 0.07 | 0.754(0.580.979) |
| Exposure | Outcome | Nsnp | Beta | SE | P value | OR (95%CI) |
|---|---|---|---|---|---|---|
| Interleukin-4 levels | Adenosine 5\'-diphosphate (ADP) to choline ratio | 10 | 0.216 | 0.098 | 0.027 | 1.241(1.0241.503) |
| Adenosine 5\'-diphosphate (ADP) to choline ratio | Hepatic cancer | 20 | 0.094 | 0.11 | 0.395 | 1.098(0.8851.364) |
| Interleukin-4 levels | Hepatic cancer | 12 | 0.309 | 0.155 | 0.046 | 1.362(1.0061.844) |
| Interleukin-4 levels | Hepatic cancer | 12 | 0.288 | 0.155 | 0.089 | 1.334(0.911.956) |
In this study, utilizing large-scale GWAS data, we conducted two-sample MR and MR mediation analyses to investigate the causal relationships between 91 inflammatory factors, 1400 serum metabolites, and hepatic cancer. After correcting for heterogeneity and excluding horizontal pleiotropy, our study confirmed significant associations between six inflammatory factors and hepatic cancer. Among these, four inflammatory factors were positively associated with an increased risk of hepatic cancer, while two inflammatory factors were negatively associated with the risk of hepatic cancer. Regarding the MR analysis results of serum metabolites and hepatic cancer, a total of 73 serum metabolites were found to be significantly associated with hepatic cancer. Among these, 41 serum metabolites were positively associated with an increased risk of hepatic cancer, while the remaining 32 serum metabolites were negatively associated with the risk of hepatic cancer. Additionally, three serum metabolites exhibited mediating effects in the influence of inflammatory factors on the risk of hepatic cancer. Inflammatory factors may influence the occurrence of hepatic cancer by modulating the levels of serum metabolites, either increasing or decreasing their levels.
In the study of the causal relationship between inflammatory factors and hepatic cancer, we found that six inflammatory factors, including C-C motif chemokine 19 levels, Fms-related tyrosine kinase 3 ligand levels, and Interleukin-13 levels, are causally related to hepatic cancer. Inflammatory factors, produced by activated inflammatory cells, comprise a class of biologically active small molecular proteins or peptides. They are typically secreted into the bloodstream, playing roles in regulating the human immune system response and maintaining physiological balance. The most common inflammatory factors include Interleukins (IL), Tumor Necrosis Factors (TNF), and Interferons (IFN). Recent studies have shown that inflammatory factors play critical roles in the occurrence and development of tumors, being considered important biological features of malignancies. Fifteen to twenty percent of malignant tumors are caused by infection and non-controllable inflammation. For instance, inflammatory bowel disease is associated with colon cancer, HBV infection with hepatic cancer, HP infection with gastric cancer, EB virus infection with nasopharyngeal cancer, and HPV virus infection with cervical cancer. The inflammatory environment is closely linked to the tumor microenvironment. Numerous inflammatory factors exist in the tumor microenvironment, such as IL-1, IL-6, IL-12, IL-17, TNF-α, and TGF-β, which not only recruit inflammatory cells to the tumor site and amplify inflammatory effects but also promote tumor cell growth and migration, as well as tumor blood vessel and lymphatic vessel formation. Specifically, IL-6 can induce microRNA-21 expression in a STAT3 signaling pathway-dependent manner, thereby promoting cell proliferation and inhibiting apoptosis. Inflammatory factors such as TNF-alpha can activate transcription factors in premalignant cells, such as NF-kB signaling, Signal Transducer and Activator of Transcription 3 (STAT3), and Activator Protein 1 (AP-1), further regulating tumor cell growth and migration, thus playing important roles in the occurrence and development of tumors. Inflammatory factors can also induce cell carcinogenesis or inhibit tumor suppressor gene mutations. Prolonged inflammatory states place cells in an oxidative stress environment, increasing the likelihood of DNA mutations and thus increasing the risk of tumor formation. TNF-α, IFN-β, and others can stimulate the expression of microRNA-155, thereby inducing tumorigenesis. Inflammatory factors are also potential tumor markers. Researchers have developed a tool to assess the inflammatory burden of cancer patients—the Inflammatory Burden Index (IBI). The IBI, a combination of C-reactive protein, neutrophils, and lymphocytes, has been experimentally validated as a potential biomarker for predicting the prognosis of cancer patients [15]. The important role of inflammatory factors in cancer makes them potential targets for tumor treatment. Currently, some drugs targeting inflammatory factors or their signaling pathways have shown promise in cancer treatment. For example, Interleukin-1 (IL-1) antagonists and Interleukin-6 (IL-6) ligand blockade antibodies have shown good efficacy in clinical trials [16]. IL-1 antagonists are being attempted for the treatment of multiple myeloma and melanoma, while IL-6 ligand blockade antibodies are currently in clinical trials for the treatment of multiple myeloma, prostate cancer, and renal cell carcinoma. Inflammatory factors also play important roles in the occurrence and development of hepatic cancer. A meta-analysis study showed that the TNF-α G-308A polymorphism increases the risk of hepatic cancer in Asians [17]. TNF-α and IL-6 promote the development of obesity-mediated hepatic cancer by promoting the development of fatty liver and steatohepatitis [18]. TNF-α can also induce the generation of AIF-1+ CSF1R+ MSCs to create an inflammatory microenvironment and promote the occurrence of hepatic cancer [19]. On the other hand, some inflammatory factors also have tumor-suppressive functions. For example, IFN-α and IFN-β can inhibit the formation of tumor blood vessels, stopping tumor growth and spread [20]. They can also enhance immune cell killing of tumors, thereby inhibiting tumor growth. Yanmeizhi Wu et al. summarized the research progress on immune checkpoint inhibitors regulating inflammatory factors to control anti-tumor responses, demonstrating that inflammatory factors play functional roles in Programmed Death-1 (PD-1) or other immune checkpoints, participating in regulating the expression of Programmed Death Ligand-1 (PDL1) and activating immune conditions to counteract tumor progression [21].
In the study of the causal relationship between serum metabolites and hepatic cancer, we found that 73 serum metabolites, including S-allylcysteine levels, (1-enyl-palmitoyl)-2-linoleoyl-GPE (p-16:0/18:2) levels, and Caffeic acid sulfate levels, are causally related to hepatic cancer. Serum metabolites are small molecular compounds produced during normal physiological and pathological processes, including proteins, carbohydrates, lipids, etc., and their concentrations reflect various biochemical processes occurring within organisms. When the body is diseased, this normal metabolic balance is disrupted. The formation of tumors is often associated with imbalances in various substance metabolisms within the body. Rapid tumor growth and energy consumption lead to a decrease in specific metabolites in the serum, while metabolites released from tumor cell death may differ from those released from normal cell death, which may be reflected in the serum. Previous studies have found that serum metabolites may be both potential causes of tumor occurrence and markers for early detection, screening, and prevention of tumors to a large extent. A category of serum metabolites has been widely used in cancer diagnosis and treatment. For example, the elevation of certain tumor markers may indicate the presence of tumors in the body. Although these tumor markers have high specificity, their sensitivity is relatively low, limiting their application in early cancer screening and diagnosis. Jesus M. Banales et al. found that changes in the concentrations of specific metabolites in the serum help distinguish intrahepatic cholangiocarcinoma from Hepatocellular Carcinoma (HCC) or primary sclerosing cholangitis and contribute to the early diagnosis of these diseases [22]. In breast cancer, changes in ER/PR and Her-2/neu are used to predict disease progression. In recent years, more and more studies have shown a close association between serum metabolites and hepatic cancer. Some metabolites have been found to be associated with the occurrence and metastasis of hepatic cancer. For example, acetylated tyrosine, glutamine, lipid metabolites, etc., exhibit abnormal levels in the serum of hepatic cancer patients, and these changes in metabolites may be closely related to the growth and metastasis of tumors [1]. Additionally, serum metabolites can also be used for monitoring and prognostic evaluation of hepatic cancer treatment, which is of great significance for formulating individualized treatment plans and improving treatment effectiveness. Ana P. Gomes’ research team found that serum Metabolite Methylmalonic Acid (MMA) can induce the expression of SOX4, leading to transcriptional reprogramming, making hepatic cancer cells aggressive, indicating that MMA is a promising target for late-stage hepatic cancer treatment [23]. Jinkai Liu et al. used a comprehensive metabolomics approach to study and found that DL-3-phenyllactic acid, L-tryptophan, glycocholic acid, and 1-methylnicotinamide in portal vein serum of HCC patients are associated with impaired liver function and poor survival, while arachidonic acid and phenol significantly inhibit hepatic cancer cell proliferation in portal vein serum of HCC patients, suggesting they are protective metabolites against hepatic cancer [24].
Our mediation analysis also provides genetic evidence for the association between inflammatory factors, blood metabolites, and hepatic cancer. To our knowledge, there has been no previous study directly linking inflammatory factors with serum metabolites. However, some studies have assessed the potential relationship between inflammatory factors and serum metabolites. Yasuyuki Shimomura et al [25]. explored the correlation between serum reactive oxygen metabolites, antioxidant markers, oxidative indices, and clinical features, finding a positive correlation between serum reactive oxygen metabolites and C-reactive protein and inflammation scores in non-alcoholic fatty liver disease, suggesting a possible positive association between inflammatory factors and serum metabolites. Serum reactive oxygen metabolites are considered reliable indicators of circulating Reactive Oxygen Species (ROS) [2,26]. Studies have reported that ROS induce the synthesis and activation of numerous cytokines and growth factors, thereby promoting the progression of hepatocellular carcinoma [27,28]. Current research results suggest that inflammation can trigger oxidative stress, which plays a crucial role in controlling adverse activities in active hepatitis. Serum reactive oxygen metabolites, specifically reduced glutathione levels, have also been shown to be negatively correlated with recognized antioxidant stress markers, indicating an association between serum metabolites and inflammation, which may affect the occurrence and development of liver diseases. Our mediation MR analysis also indicates that the inflammatory factor Interleukin-13 levels and Interleukin-4 levels can respectively increase the levels of serum metabolites Linolenate [alpha or gamma; (18:3n3 or 6)] levels and Adenosine 5’-Diphosphate (ADP) to choline ratio, thereby increasing the risk of hepatic cancer occurrence, while the inflammatory factor Monocyte chemoattractant protein-3 levels can decrease the levels of serum metabolite Sphingomyelin (d18:0/20:0, d16:0/22:0), thereby reducing the risk of hepatic cancer occurrence. Thus, some of the effects of inflammatory factors on the risk of hepatic cancer occurrence are achieved through influencing the levels of serum metabolites.
This study conducted dual-sample MR and MR mediation analyses based on published results from large GWAS cohorts, which have several notable advantages. Firstly, this study employed genetic instrumental variables and utilized various MR analyses to determine the causal relationships between inflammatory factors, serum metabolites, and hepatic cancer, thereby minimizing the influence of confounding factors and reverse causation as much as possible. Secondly, compared to prospective cohort studies or randomized controlled trials, MR analysis allows easier access to publicly available data, reducing the time and cost required for the study. Thirdly, the study had a large sample size of European populations, thus possessing higher statistical power. Lastly, given the association between inflammatory factors and hepatic cancer development, our study further complements existing evidence and reveals the mediating role of serum metabolites in its development, potentially providing insights for clinical diagnosis and treatment. Nevertheless, our study still has limitations. Firstly, despite conducting multiple sensitivity analyses, we could not fully evaluate horizontal pleiotropy. Secondly, the study was based on European databases, and the conclusions may not be generalizable to other ethnic groups, warranting further research for applicability to other ethnic populations. Thirdly, to include more inflammatory factors in the MR analysis, we used a looser threshold to assess the results, which may increase some false positives. Additionally, we could not ascertain the overlap between the three large sample datasets. Finally, our conclusions need to be validated through high-evidence randomized controlled trials.
In conclusion, this study utilized publicly available GWAS data to conduct MR analysis on the causal relationships between inflammatory factors, serum metabolites, and hepatic cancer. We ultimately identified causal relationships between 6 inflammatory factors and hepatic cancer, as well as 73 serum metabolites and hepatic cancer. Among them, 5 inflammatory factors were significantly associated with hepatic cancer, along with 73 serum metabolites. Mediation MR analysis indicated that three serum metabolites mediate the effect of inflammatory factors on the risk of hepatic cancer incidence. Inflammatory factors may influence the risk of hepatic cancer occurrence by altering the levels of serum metabolites. This finding provides a new perspective for further understanding the role of inflammatory factors in the development of hepatic cancer and offers potential targets for the prevention and treatment of related diseases.
Provide a data availability statement:The datasets generated and analyzed during the current study are available in the catalog database (https://www.ebi.ac.uk/gwas/publica tions/37563310).
Declaration of interest statement:The authors unequivocally state that they possess no known competing financial interests or personal relationships that could have conceivably influenced the integrity or outcomes of the work presented in this paper.
Acknowledgment:This study was supported by the National Natural Science Foundation of China (82205224), the Hunan Provincial Institute of Traditional Chinese Medicine Key Project (202116), the Hunan Provincial Graduate Research and Innovation Project (CX20230835).
Author contributions statement:All authors have had access to the data and all drafts of the manuscript. Specific contributions are as follows: data collection, data management, data analysis, and manuscript drafting: Weixin Wang, Linpei Li, Bicai Yang, Siyu Shen, Wenchun Chen, Hongyao Chen. Study design: Weixin Wang. Data analysis and chart visualization: Linpei Li. Manuscript review: all. Funding sources: Bo Pan.